|
001 ИСТОРИЯ эволюции человечества в версии НАУКИ
|
|
| kualspb_2013 | Дата: Пятница, 15.11.2019, 18:43 | Сообщение # 1 |
 Генералиссимус
Группа: Администраторы
Сообщений: 2147
Статус: Offline
| ИСТОРИЯ эволюции человечества в версии НАУКИ
-- Приведу здесь версию ученых о том, как развивалась жизнь на планете Земля. Этот же вариант размещен мною внутри темы, где дается версия Махатм через Елену Петровну Блаватскую.
_001
В Млечный путь входит Солнечная система. Она включает в себя Солнце – звезду, масса которой в 109 раз больше массы Земли. От Солнца до центра Галактики около 30 тысяч световых лет. Период обращения Солнца вокруг центра галактики около 250 миллионов лет.
Солнечная система – это 9 планет, 40 тысяч астероидов, вращающихся по различным орбитам вокруг Солнца. Солнечная система образовалась много позднее метагалактики, всего 4,5–5 млрд. лет назад. Расстояние от Солнца до самой далекой планеты Плутона около 6 млрд. километров. 99,86% всей массы Солнечной системы сосредоточено в самом Солнце. Солнце представляет собой раскаленный плазменный шар. Его температура на видимой поверхности колеблется между 5000 и 6000 0С, а температура ядра, где идут ядерные реакции и вырабатывается солнечная энергия, достигает 15 000 000 0С. По химическому составу Солнце состоит из 90% водорода, 10% гелия.
Земля – третья планета по удалению от Солнца. Для удобства принято считать ее массу в Солнечной системе равной 1, а массы остальных небесных тел солнечной системы рассчитывать по сравнению с ней. Средняя температура поверхности Земли +12 ◦С, атмосферное давление 1 атм., ускорение свободного падения на экваторе 981 см/сек2, среднее расстояние до Солнца 1 астрономическая единица (149,6 млн. км), радиус 6,96 108 м.
Поверхность Земли более чем на 2/3 покрыта океаном (70,8%), средняя высота суши – 875 м над уровнем моря, средняя глубина океана – 3800 метров. Самое высокое место нашей планеты – гора Джомолунгма (Эверест), ее высота составляет 8848 м, самое глубокое место в океане – Мариинская впадина, чья глубина – 11022 м.
В атмосфере Земли по объемному содержанию приблизительно 78% азота, 21% кислорода, а на оставшийся 1% приходятся углекислый газ, метан, инертные и иные газы. Этот состав установился в процессе эволюции на Земле. Кислород появился в составе атмосферы 2-2,5 миллиарда лет назад в результате фотосинтеза, осуществляемого различными видами растительных организмов, имеющими в своем составе хлорофилл.
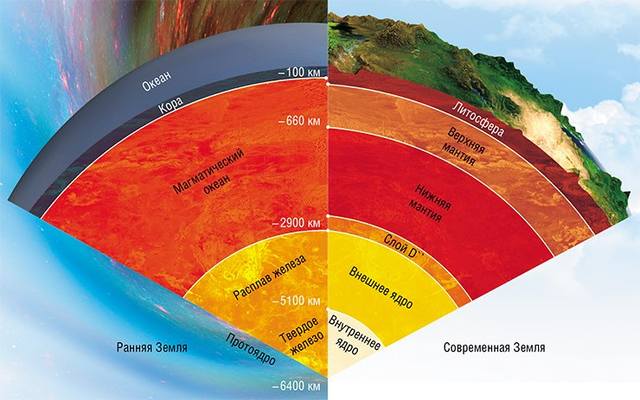
В центре Земли на глубине 6378 км находится жесткое ядро, состоящее из железо-никелиевого сплава. Ученые полагают, что оно покрыто оболочкой жидкого оксида железа Fe2О, толщиной около 1200 км. Затем следует переходный слой и мантия, простирающаяся от 2900 до 900 км от поверхности Земли, и земная кора, состоящая до 70 км от поверхности из базальтового слоя, до 40 км – из гранитного слоя и далее до 20 км осадочного слоя.
 https://u.to/eQNUFg https://u.to/eQNUFg
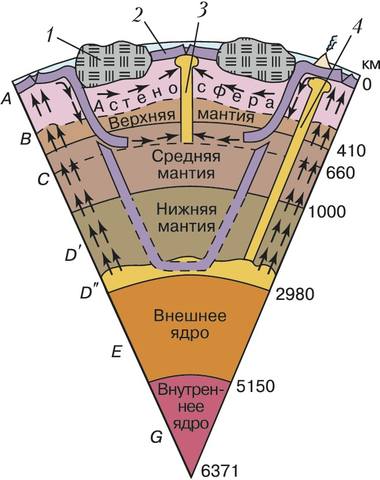
Схема внутреннего строения и динамики твёрдой Земли (А – земная кора и литосферная мантия; стрелками показано направление переноса вещества): 1 – континентальная литосфера; 2 – океаническая литосфера; 3 – плюм исландского типа; 4 – плюм гавайского типа.
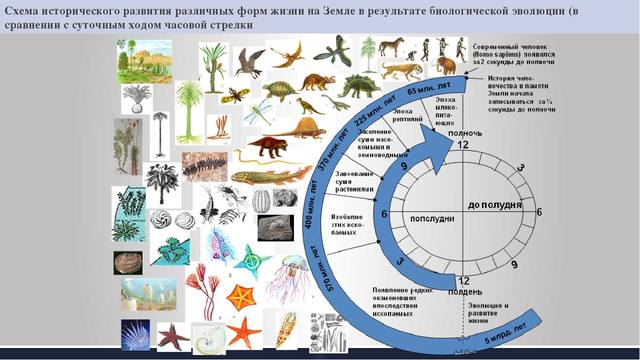
Если принять историю Земли за год, то Земля образовалась 1 января, 12 декабря – время расцвета динозавров. И только 31 декабря появляется общий предок обезьяны и человека.
 х х 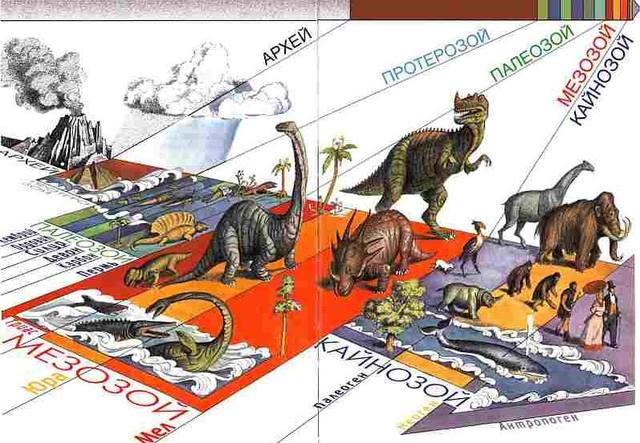
Любопытно, что Земля «стала» древней планетой меньше века назад. Только в 1913 году Артур Холмс, британский геолог, опубликовал книгу «Возраст Земли». Он предложил использовать для датировки горных пород уран-свинцовый метод – по соотношению между ураном (распадающимся до свинца) и свинцом в породе, как песку в песочных часах.
С этого момента стали говорить о возрасте Земли в миллиардах, а не в миллионах лет. К настоящему времени в Австралии найдены кристаллы циркона с возрастом порядка 4,4 млрд. лет. Эти кристаллы также показывают, что уже в то время на Земле существовала водная оболочка.
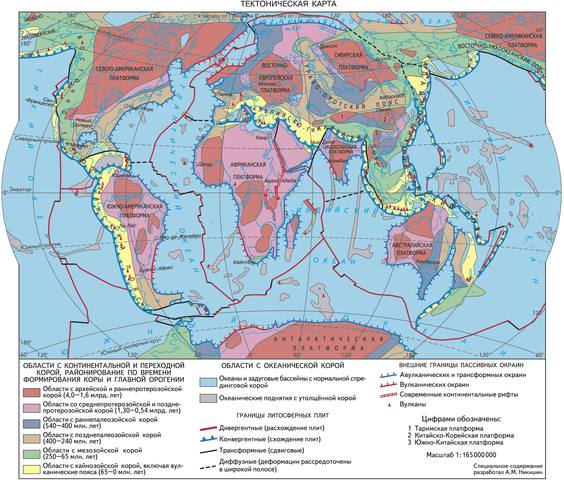
Фиксируя сейсмические толчки, используя системы спутниковой навигации и компьютерного анализа (позволяющие быстро обрабатывать большие массивы информации), ученые установили, что земная кора находится в постоянном движении. Многие миллионы лет назад участки земной коры были в совершенно других местах, чем мы привыкли их видеть.
В 1950-х гг. английский физик Патрик Блэкетт и геофизик Кейт Ранкорн независимо друг от друга изучали остаточную намагниченность различных пород во многих уголках Земли - в Англии, Индии, Исландии, Африке, США, Антарктиде. Дело в том, что при формировании некоторых горных пород содержащиеся в них ферромагнитные частицы ориентируются по направлению магнитного поля Земли, сохраняя направление на магнитный полюс. Многократные измерения показали, что направление на магнитный полюс в древних породах не совпадает с современным, и чем древнее породы, тем расхождение больше. Конечно, магнитные полюса могли бы менять свое положение, но для разных регионов их картина не получалась единой – все одновозрастные породы с одного континента указывали на один и тот же полюс, но образцы с разных континентов давали и разные положения полюсов. В таком случае, перемещались сами породы, т. е. континенты?
Одной их нераскрытых тайн прошлого является возникновение жизни на Земле. Само определение жизни, столь очевидное, поскольку на наших глазах жизнь проявляется многогранно, не так просто. Жизнь — это форма существования организмов, возможная только в открытых системах, подпитываемых из внешнего источника энергией. Живые организмы характеризуются тем, что они в состоянии обмениваться постоянно или хотя бы периодически друг с другом и средой своего обитания энергией, веществом и информацией. Все организмы, известные нам, построены на базе химических соединений, имеющих в своем составе углерод, кислород, водород, азот, фосфор и серу. Они состоят из клеток. Наследственная информация записана единым кодом в виде совокупности нуклеиновых кислот, расположенных в определенном порядке в геноме.
Любая форма живого вещества является неизмеримо более сложной системой по сравнению с любой формой косной (неживой) материи на нашей планете. Чтобы возникла и могла поддерживать свои функции такая сложная структура, необходим приток энергии. Энергия — неотъемлемое свойство движения материи. Это способность тела, вещества, системы или любых других форм материи производить работу или теплообмен между объектами с различной температурой. В соответствии со вторым законом термодинамики в любой системе происходит выравнивание энергии, то есть все упорядоченные структуры стремятся перейти к состоянию беспорядка, хаоса.
Большинство законов, изучаемых в школе, относится к описанию замкнутых систем. В замкнутой системе ни возникновение жизни, ни ее поддержание принципиально невозможно. Это может происходить в так называемых открытых системах, где степень упорядоченности в одном месте происходит за счет увеличения беспорядков в другом. Жизнь возможна только в открытой системе.
Ученые полагают, что в первобытном океане примерно 3,5 млрд. лет назад произошли соединения сложных органических веществ в новые еще более сложные субстанции. У некоторых из них обнаружилось поразительное свойство – способность к воспроизведению своей структуры. Из окружавшего их «первичного бульона» они выбирали полезные для них вещества, тем самым поддерживая свою жизнедеятельность, в частности, рост и размножение. Об этих первых организмах мы знаем очень мало. Они, несомненно, были микроскопических размеров, подобно вирусам, бактериям или грибам. Кислород для них был ядом. Необходимая им энергия получалась в процессе ферментации веществ, входящих в состав «первичного бульона».
Затем, через 500 млн. лет, произошло событие, обеспечившее развитие жизни, – возникновение хлорофилла. Стал возможен процесс фотосинтеза, при котором из углекислого газа (существующего в окружающей среде того периода благодаря происходившим процессам ферментации) и воды на свету образуются углеводы и кислород. Появился независимый от первичного бульона источник питания.
Первые 3 млрд. лет существования жизни были исключительно бедны с точки зрения биоразнообразия. Оно было представлено главным образом одноклеточными бактериями и примитивными водорослями. Именно они определяли облик жизни на Земле. Этот период охватывает 85% всей истории существования жизни на Земле.
Почти все виды растений сложились в последние 600–400 млн лет, затем растительность начала бурно развиваться во всем ее многообразии. Появились первые животные. После выхода растений на сушу началось ее освоение. К новым условиям приспособились некоторые рыбы. Появились амфибии. Поразительна эволюционная изменчивость всех форм жизни, все более гармонично вписывающихся в условия меняющейся окружающей среды. Это определялось тем, что в популяции у разных особей одного вида имело место некоторое генетическое разнообразие, гены не были идентичны. Идею эволюции жизни развил Ч. Дарвин (1809-1882) в 1858 г. Он сформулировал ее как «естественный отбор в результате выживания наиболее приспособленных» особей вида. Идея не раскрывала механизма эволюции. Он стал понятным много позднее с развитием генетики и обнаружением генетического разнообразия организмов, входящих в одну популяцию.
Способность к эволюции неодинакова у разных видов. В большой степени она зависит от продолжительности жизни, то есть от частоты смены поколений. Поэтому бактерии, насекомые, грызуны и другие организмы, имеющие короткий период воспроизводства, приспосабливаются к меняющимся условиям весьма быстро. Примером этого стало появление устойчивости болезнетворных бактерий к пенициллину, насекомых к пестицидам и пр. И наоборот, виды, имеющие длительный период воспроизводства и малочисленное потомство, требуют миллионов лет для обнаружения у них каких-либо изменений. Так современный человек генетически мало отличается от кроманьонца.
В соответствии с летописью окаменелостей, 94–99% видов живых существ, живших на Земле, вымерли к настоящему времени. Сейчас количество видов оценивается 5– 30 миллионами, причем ежегодно описывается около тысячи неизвестных видов.
Высшим проявлением эволюции на Земле стал человек разумный (Homo sapiens). Организм любого человека – самое сложное сочетание молекул в рамках одного организма на Земле, а может быть и во всей Вселенной. История человека прослеживается до некоторых приматов, существовавших 5–6 млн. лет назад. За время этой длинной, а в геологическом смысле очень короткой, эволюции человек стал, по выражению Уильяма Шекспира (1564-1616), «венцом всего живущего».
Мозг человека в результате эволюции изменялся одновременно с изменениями других органов. Три важнейшие характеристики отличают человека от животных. Его скелет приспособлен для прямохождения. Его глаза обеспечивают резкое, объемное и цветовое восприятие окружающего мира. Его руки способны осуществить как силовой хват, так и тончайшие манипуляции. Кроме того, кожа человека способна к осязанию.
В результате развития всех этих способностей у человека сформировалась речь, возможность воспринимать и передавать информацию от одного человека к другому, накапливать и использовать коллективный опыт. История становления человека по оценкам антропологов составляет менее 3 млн. лет. Это очень короткий период в истории жизни на Земле. Если представить время с момента появления жизни на земле, как суточный циферблат часов, современный человек появился всего за 2 секунды до полуночи, а его история начала записываться за 1/4 секунды. Развитие человечества протекало лишь в последние 2 секунды этих условных суток, но, несмотря на столь кратковременную жизнь, мы сейчас становимся свидетелями вымирания большего числа земных видов, чем за предыдущие 5 млрд. лет.
(ЩЕЛКНИТЕ ПО КАРТИНКЕ, ЧТОБЫ ЕЕ УВЕЛИЧИТЬ)
 х х 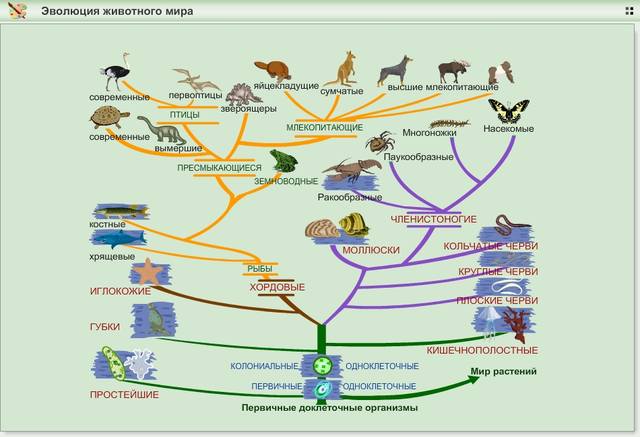
 х х 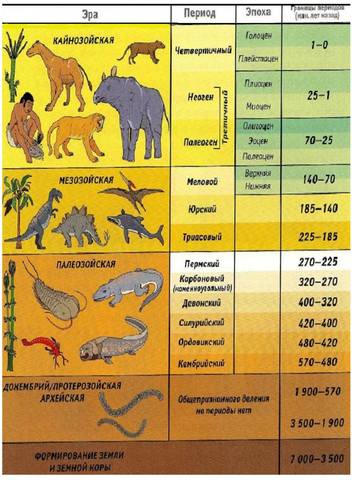
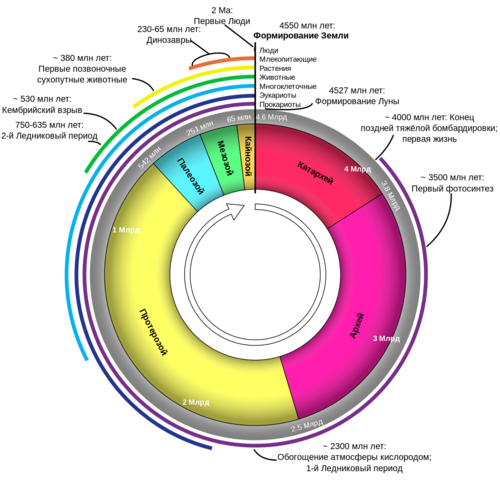 х х 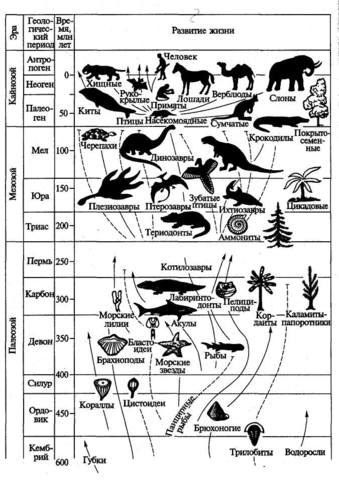
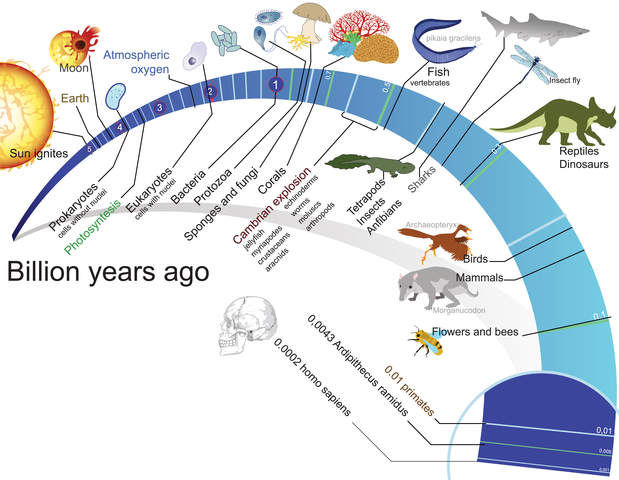 х х 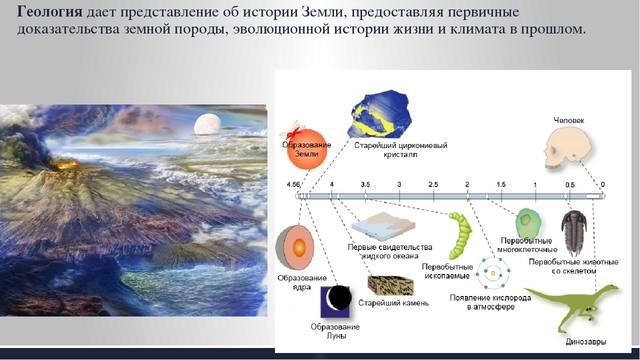
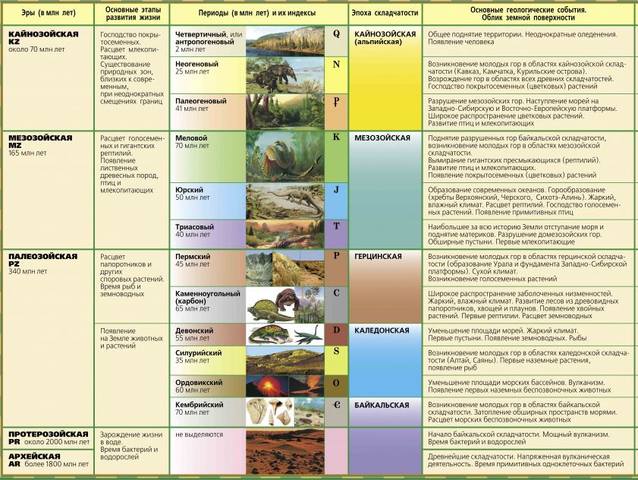
Как исчисляется возраст Земли. При изучении возраста Земли составили календарь Земли. История Земли разделена на длительные промежутки времени – эры. Эры делятся на периоды, периоды на эпохи, эпохи – на века. Названия эр греческого происхождения: архейская – древнейшая, протерозойская – ранняя, палеозойская – древняя, мезозойская- средняя, кайнозойская – новая. На основе определения геологического возраста горных пород учёные составляют геохронологические таблицы. Чтение таких таблиц начинают снизу по мере залегания горных пород.
 х х 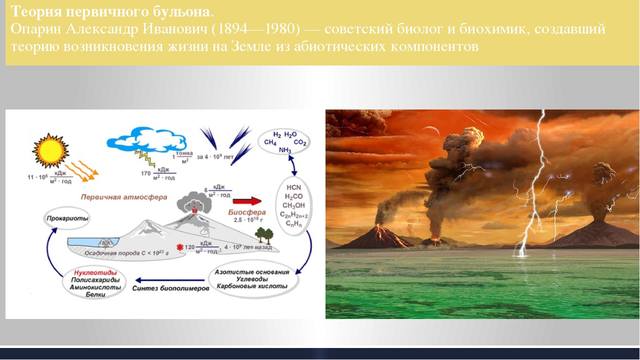
 х х 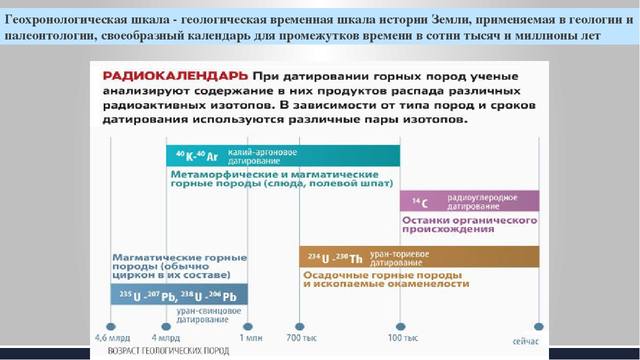
ххххххххх
|
| |
|
|
| kualspb_2013 | Дата: Пятница, 15.11.2019, 18:48 | Сообщение # 2 |
|
Генералиссимус
Группа: Администраторы
Сообщений: 2147
Статус: Offline
| _002
Согласно расчетам Л. Беркнера и Л. Маршалла, освоение организмами суши как среды обитания могло начаться во второй половине ордовикского периода, когда содержание кислорода в земной атмосфере достигло 0,1 от современного. Действительно, первые вполне достоверные остатки наземных организмов известны начиная с силура. Вероятно, заселение безжизненных прежде материков было длительным процессом, развивавшимся в течение нескольких десятков миллионов лет, на протяжении ордовика, силура и девона.Пионерами на суше были, несомненно, растения, сначала заселившие мелководья у морских побережий и пресные водоемы, а затем постепенно осваивавшие влажные местообитания на берегах. Древнейшими представителями этой «земноводной» флоры были псилофиты (Psilophyta) — невысокие травянистые или кустарникообразные растения (рис. 25), еще не имевшие настоящих корней, с мелкими листочками и органами бесполого размножения (спорангиями) на концах дихотомически разветвленных побегов. В их древесине уже имелись примитивные сосудистые образования — трахеиды. Псилофиты известны начиная с позднего силура и достигли расцвета в раннем и среднем девоне. Заселение суши растениями положило начало почвообразованию с обогащением минеральных субстратов органическими веществами. К этому времени (силур — девон) относится и возникновение наиболее древних толщ горючих ископаемых, образующихся при неполном разложении скоплений растительных остатков.
Во влажном и теплом климате, характерном для первой половины каменноугольного периода, широкое распространение получила обильная наземная флора, имевшая характер густых влажных тропических лесов
По мере заселения суши растениями появились предпосылки для освоения наземной среды обитания животными. Скорее всего, первыми среди них были какие-то мелкие растительноядные формы, существовавшие в силурийском периоде, но не оставившие следов в палеонтологической летописи (во всяком случае, их остатков пока не обнаружено). Как отметил М. С. Гиляров (1971), для многих групп беспозвоночных животных освоение суши началось с использования почвы как промежуточной среды обитания, в которой многие условия (повышенная влажность, степень плотности субстрата) были в какой-то мере близки к таковым водной среды
Вероятно, освоение суши животными началось уже в раннем силуре, хотя из силура пока почти неизвестно ископаемых остатков наземных животных, за исключением скорпиона Palaeophonus из силурийских отложений Англии 1. Из девона известны представители уже нескольких групп наземных членистоногих: палеозойская группа панцирных пауков (Soluta), клещи (Acaromorpha) и низшие первичнобескрылые насекомые (Apterygota). Несомненно, разнообразие наземных беспозвоночных в девонском периоде было значительно большим, чем это представляется по дошедшим до нас скудным ископаемым остаткам. Так, многоножки (примитивнейшая группа трахейных членистоногих) известны лишь начиная с позднего карбона, но, несомненно, они существовали уже в первой половине девона. Во второй половине раннекарбоновой эпохи появились наделенные крыльями высшие насекомые, принадлежащие к подклассу Pterygota. Вероятно, предки крылатых насекомых перешли к лазающей жизни на стволах и в кронах деревьев. Для облегчения планирования при падениях и прыжках у этих животных служили боковые уплощенные выступы стенки тела на грудных сегментах. Из этих неподвижных придатков в процессе длительной приспособительной эволюции естественный отбор сформировал удивительный в своем совершенстве летательный аппарат.
Вслед за беспозвоночными к освоению наземной среды обитания приступили и позвоночные. Их выход на сушу мог осуществиться лишь после появления там соответствующей пищевой базы (достаточно обильной фауны наземных беспозвоночных).
В карбоне начинается расцвет древних амфибий, представленных в позднем палеозое большим разнообразием форм (рис. 29), которых условно объединяют под названием стегоцефалов, т. е. «покрытоголовых».
От каких-то примитивных батрахозавров (пока еще неизвестных) в раннем карбоне возникли настоящие рептилии (Reptilia)
Одновременно у древнейших рептилий возник второй комплекс приспособлений, избавивших от связи с водоемами все важнейшие процессы, связанные с размножением. Это было появление внутреннего осеменения вместо характерного для земноводных наружного; формирование крупных яиц, обильно снабженных запасными питательными веществами (желтком) и способных обеспечить длительное развитие зародыша; образование плотных яйцевых оболочек, защищающих яйцо на суше от обезвоживания и механических повреждений
Животный мир также претерпел значительные изменения на протяжении пермского периода, ставшие особенно драматическими во второй его половине. В наибольшей степени это коснулось морских животных, среди которых многие группы подверглись значительному угнетению, сопровождавшемуся уменьшением численности и разнообразия видов, вплоть до полного вымирания целых классов. Примерно в середине пермского периода вымирают последние трилобиты, эвриптериды, бластоидеи (один из классов прикрепленных иглокожих), палеозойские группы морских лилий. К концу перми исчезают тетракораллы, значительно сокращается число видов плеченогих, мшанок, морских ежей, офиур, аммоноидей и наутилоидей среди моллюсков, остракод среди ракообразных, губок, фораминифер среди простейших. Из позвоночных вымирают акантодии, многие палеозойские группы хрящевых рыб. В пресных внутриконтинентальных водоемах значительно снижается численность хоановых рыб. К концу палеозоя вымирают лепоспондильные стегоцефалы. Изменения в наземной фауне протекали в целом не столь драматично, хотя и здесь произошли существенные изменения. Пермское вымирание по масштабам принадлежит к категории так называемых «великих вымираний» и сравнимо, например, с широко известным великим вымиранием в конце мелового периода. Причины великих вымираний, приводивших к радикальным изменениям общего облика фауны и флоры, в большой степени еще остаются загадочными.
ГЛАВА 4. МЕЗОЗОЙСКАЯ ЭРА — ВЕК РЕПТИЛИЙ
После завершения в середине пермского периода оледенения Гондваны климат Земли стал более теплым. (Возможно, это было связано с перемещением Южного полюса с континента в Тихий океан.) Потепление продолжалось на протяжении мезозойской эры, которая в целом была более однообразной в климатическом отношении, чем другие эры фанерозоя. В мезозое господствовали теплые климаты с относительно слабо выраженной климатической зональностью.
Зато появившиеся в позднеюрское время крупные плиозавры (Pliosauroidea) имели огромный череп (до 3 м у 12-метровых ящеров) с острыми зубами длиной до 10 см. Эти морские хищники могли, вероятно, питаться крупными глубоководными головоногими (подобно современным кашалотам) и даже нападать на более мелких плезиозавров и других морских рептилий.
ВЕК ДИНОЗАВРОВ
К концу триаса произошло постепенное «выравнивание» климатических условий на значительной части континентов, о чем говорит, в частности, очень однообразный и в целом сходный характер флоры на больших пространствах континентов, входивших прежде в состав Гондваны и Лавразии. Растительность юрского периода (рис. 46) характеризовалась преобладанием различных групп голосеменных (цикадовые, беннеттиты, хвойные, гинкговые) и древовидных папоротников; широко распространены были также крупные хвощи (среди которых, например, Equisetites arenaceus достигал в высоту 10 м и в диаметре 25 см). В условиях ровного и очень теплого климата в средней части мезозоя завропсидные рептилии достигли максимального расцвета. Это было время удивительных животных, получивших широкую известность под названием «динозавры» («страшные ящеры»).
В юрском и меловом периодах динозавры доминировали среди наземных животных, заселив всевозможные местообитания и дав огромное разнообразие форм, которые поражают воображение гротескной причудливостью облика, а иногда и огромными размерами. Безусловный рекорд в последнем отношении принадлежит ящеротазовым динозаврам из группы завропод (Sauropoda), среди которых были самые крупные из когда-либо существовавших на Земле четвероногих животных. Например, диплодок (Diplodocus, рис. 48) достигал в длину 25—30 м при высоте в области середины спины около 4 м, масса этого животного составляла около 30 т. Несколько более короткий (длиной 24 м) брахиозавр (Brachiosaurus) имел более высокие передние конечности и массивное тело, масса которого, по расчетам, могла достигать 45—50 т.
Правда, и завроподы значительную часть жизни, скорее всего, проводили в воде или по берегам водоемов. Об этом говорят, помимо гигантских размеров и массы, при которых очень трудно свободно передвигаться по суше, многие особенности строения этих удивительных животных
Второй группой теропод были карнозавры (Carnosauria), к которым принадлежали самые крупные хищники, когда-либо существовавшие на суше (мегалозавры — Megalosauridae и дейнодонты — Deinodontidae). Некоторые из этих чудовищ, например тираннозавр, или «царственный ящер — тиран» (Tyrannosaurus rex, рис. 50), достигали в длину 12—14 м. Полутораметровый череп, вооруженный острыми, кинжалообразно изогнутыми зубами с пильчатыми краями (длиной до 10—15 см), находился на высоте 4—6 м. По расчетам, масса тираннозавров достигала 2 т.
Как и у современных рептилий, головной мозг динозавров имел относительно небольшие размеры. Особенно это бросается в глаза у гигантских форм, полость мозговой коробки которых поражает своими ничтожно малыми размерами по отношению к размерам черепа и всего тела. Зато спинной мозг был сильно утолщен в крестцовой области, причем нередко это утолщение было много больше по объему, чем головной мозг. Вероятно, здесь находились нервные центры, ответственные за работу задних конечностей, хвоста и за различные безусловнорефлекторные реакции. Особенно велико было крестцовое утолщение спинного мозга у завропод.
Архозавры в мезозое овладели не только сушей, но и воздухом (их попытки выйти в море были менее успешны). По крайней мере две группы этих рептилий приобрели способность к полету.
Долгое время рекорд размеров принадлежалпозднемеловомуптеранодону (Pteranodon) — огромному ящеру, размах крыльев которого составлял 7,5—8 м, а их площадь — 5,8 м2; масса тела птеранодона достигала 18—25 кг.
Птерозавры были единственными летающими позвоночными на протяжении большей части юрского периода. В поздней юре у них появились конкуренты, дальнейшие эволюционные преобразования которых привели к формированию более совершенного летательного аппарата и к достижению более высокого общего уровня организации. Это были птицы.
ИЗМЕНЕНИЯ В СОСТАВЕ НАЗЕМНЫХ БИОЦЕНОЗОВ ВО ВТОРОЙ ПОЛОВИНЕ МЕЗОЗОЯ
На фоне поражающей воображение эволюции архозавров, во второй половине мезозоя происходили и многие другие, может быть, внешне менее эффектные, но столь же интересные и важные события. В начале мелового периода от ящериц возникли змеи (Ophidia).
Млекопитающие в течение «века динозавров» оставались сравнительно малозаметными членами мезозойских биоценозов. Это были небольшие зверьки, которые вели скрытный образ жизни, избегая открытых пространств (вероятно, большинство из них имело сумеречную или ночную активность, см. выше). Однако и среди млекопитающих происходила адаптивная радиация. Помимо разнообразных насекомоядных и мелких хищников — триконодонтов (Triconodontia), докодонтов (Docodontia), симметродонтов (Symmetrodontia), пантотериев (Pantotheria), уже в конце триаса появились и растительноядные звери, принадлежавшие к отряду многобугорчатых (Multituberculata)
Может быть, наиболее значительным по своим последствиям событием, происшедшим примерно в середине «века динозавров» и значительно изменившим облик позднемезозойских биоценозов, было появление покрытосеменных, или цветковых, растений (Angiospermae). Первые достоверные остатки представителей этой группы высших растений встречаются в нижнемеловых отложениях (с абсолютным возрастом около 120 млн. лет). Древнейшие покрытосеменные были мелколистными кустарниками или небольшими деревьями. В начале позднего мела покрытосеменные приобретают самое широкое распространение и огромное разнообразие размеров и форм. Среди них становятся многочисленными крупнолистные растения, в том числе представители современных семейств магнолиевых, лавровых, платановых и др.
Но в конце мелового периода в относительно краткие (геологически) сроки произошло вымирание многих групп позвоночных и беспозвоночных животных, наземных, водных и летающих организмов. Вымирают и гигантские формы, и животные мелких размеров, и растительноядные, и хищные. К началу кайнозоя вымерли все динозавры, 8 из 10 верхнемеловых семейств крокодилов, все летающие ящеры, плезиозавры, мозазавры, последние виды ихтиозавров. Среди беспозвоночных вымирание постигло широко распространенных в юрском и меловом периодах двустворчатых моллюсков-рудистов, аммонитов, белемнитов и многих наутилоидных головоногих, вымерли также многие виды морских лилий и фораминифер. Следует подчеркнуть, что это великое вымирание не сопровождалось одновременным повышением численности и разнообразия видов каких-то других групп. Как и в пермском периоде, произошло значительное общее обеднение фауны. Уже в кайнозое (так сказать, на освободившемся месте) начинается экспансия не затронутых вымиранием групп (млекопитающие, птицы, наземные лепидозавры, бесхвостые земноводные). С другой стороны, как и во время пермского вымирания, на рубеже мезозоя и кайнозоя некоторые группы животных как бы остались в стороне от происходивших событий (их разнообразие и численность не претерпели существенных изменений). Среди позвоночных это различные группы рыб, хвостатые земноводные, черепахи. Как и в пермском периоде, великое вымирание в конце мела не носило характера мировой катастрофы (физико-географические условия на рубеже мезозоя и кайнозоя не претерпели каких-либо внезапных и резких изменений) и сам процесс вымирания был кратким лишь в геологическом смысле. Он продолжался в течение миллионов лет, когда вымирающие филетические линии постепенно угасали.
Поскольку подвергшиеся вымиранию группы организмов в конечном итоге исчезли повсеместно, многие ученые предполагали, что причины таких явлений должны были иметь характер всемирных катастроф. Первая из «катастрофических» гипотез была выдвинута еще Ж. Кювье, который считал причиной великого вымирания в конце мела вулканическую деятельность, связанную с альпийской фазой горообразования.
Однако в фанерозое проявления вулканизма всегда имели локальный характер, и непосредственный эффект вулканической деятельности мог сказываться лишь на относительно небольшой части земной поверхности. С другой стороны, горообразовательные процессы, сопровождавшиеся вулканизмом, происходили в разных регионах земного шара и в юрское время, и в меловом периоде задолго до эпохи великого вымирания (древне- и новокиммерийские и австрийская фазы альпийского горообразования, см. выше), не приводя к катастрофическим последствиям для мира динозавров и их современников.
Высказывались предположения о связи вымирания динозавров с изменениями биотических факторов, в качестве которых называли, в частности, конкуренцию со стороны млекопитающих и преобразования флоры, связанные с широким распространением покрытосеменных в середине мелового периода. Однако млекопитающие возникли еще в позднем триасе и на протяжении примерно 130 млн. лет, прошедших до конца мезозоя, оставались малозаметной и малозначительной группой животных.
Мезозой был в целом эрой низкого стояния материков. Альпийская фаза горообразования, постепенно развивавшаяся в юрском и меловом периодах, сопровождалась значительным общим поднятием материков к концу мезозоя. Результатом этого (а также снижения прозрачности атмосферы вследствие вулканической деятельности) было, по Д. Аксельроду и Г. Бейли, постепенное понижение среднегодовой температуры в течение 20 млн. лет примерно на 5°. Но, вероятно, еще более существенным фактором было возрастание неравномерности температурных условий в умеренном поясе, с развитием все более резко выраженной сезонности климата и значительным возрастанием перепада между максимальными и минимальными температурами.
Таким образом, вымирание как наземных, так и морских групп организмов в конце мелового периода могло быть в конечном итоге следствием горообразовательных процессов и поднятия континентов во второй половине мезозоя. Эти постепенно происходившие преобразования земной поверхности медленно, но неуклонно нарастали, приводя к изменениям морских течений и преобладающих направлений атмосферных потоков. Воздействие этих изменений на флору и фауну было чрезвычайно сложным и многообразным, как непосредственным, так и косвенным. Изменения условий существования, вызвавшие упадок и вымирание определенных видов и групп организмов, неизбежно должны были привести к нарушению равновесия в природных сообществах организмов — биоценозах, в которых разные виды живых существ связаны тесной зависимостью. Биоценозы обладают определенной устойчивостью, способностью к саморегуляции, но устойчивость эта имеет свои пределы. Как мы уже упоминали, если нарушения структуры биоценоза выходят за эти пределы, начинается распад всей экосистемы. При этом нарушаются сложившиеся пути передачи органических веществ и энергии в биосфере. Тогда вымиранию могут подвергнуться новые виды, которые раньше были мало затронуты непосредственно изменениями абиотических факторов. Этот процесс будет нарастать, напоминая цепную реакцию, пока тем или иным путем не будет достигнуто новое равновесие между биосинтезом и потреблением, между видами растений, растительноядных животных, хищников и микроорганизмов, т. е. пока не сформируются новые устойчивые и способные к саморегуляции экосистемы — биоценозы.
ххххххххх
|
| |
|
|
| kualspb_2013 | Дата: Пятница, 15.11.2019, 18:49 | Сообщение # 3 |
|
Генералиссимус
Группа: Администраторы
Сообщений: 2147
Статус: Offline
| _003
ГЛАВА 5. КАЙНОЗОЙ — ВЕК МЛЕКОПИТАЮЩИХ
Итак, физико-географические условия в начале палеогена, по существу, не отличались от таковых в конце мелового периода. После некоторого похолодания климат вновь стал теплым. В Европе до берегов Балтики продолжали существовать тропические и субтропические флоры, имевшие характер густых и влажных лесов. В них были широко представлены пальмы, вечнозеленые лавры, дубы, каштаны, магнолии, мирты, фикусы и другие растения; из хвойных — гигантские секвойи, араукарии, болотные кипарисы; в лесной тени произрастали папоротники, в том числе древовидные; по морским побережьям — тропическая мангровая растительность (А. Н. Криштофович, 1957). Тропические и субтропические флоры преобладали в начале палеогена также в Северной и Южной Америке, Африке и Южной Азии. Даже в Гренландии и на Шпицбергене была разнообразная растительность, свойственная теплому умеренному климату: секвойи, болотные кипарисы, гинкго, буки, дубы, платаны, клены, магнолии, липы, березы и другие растения. Любопытно развитие у многих растений высоких широт гигантских листьев (до 30—40 см). Климат в этих местах в палеогене был близок к современному климату Южной Франции или Калифорнии.
Во второй половине палеогена началось постепенное глобальное похолодание климата, которое прогрессировало в неогене. Оно завершилось в плейстоценовой эпохе четвертичного периодаразвитием нового великого оледенения. Процесс похолодания климата был весьма медленным и постепенным. Все более отчетливо проявлялась климатическая зональность. Климатические изменения легко прослеживаются по преобразованиям флоры. Теплолюбивая флора в Евразии и Северной Америке постепенно отступала к югу, сменяясь в умеренной зоне листопадными и хвойными лесами.
Согласно представлениям теории тектоники литосферных плит (см. выше), в кайнозое продолжалось движение континентов, которое постепенно привело к современной картине их распределения. От Северной Америки отделилась Гренландия. Смещения Африки и Индии почти совсем закрыли мезозойский океан Тетис. (На его месте ныне существуют Средиземное, Черное и Каспийское моря, а на востоке — моря Зондского архипелага, но среди ученых еще нет единства мнений в проблеме преемственности этих бассейнов по отношению к Тетису). Индостанская плита, смещаясь к северо-востоку, наконец, соединилась с Азией. Австралия, вероятно, лишь в эоцене обособилась от Антарктиды и стала быстро (в геологическом смысле) продвигаться к северу, приближаясь к экватору (рис. 39). Смыкание континентальных плит сопровождалось развитием горной складчатости в пограничных зонах. В кайнозое этому отвечает развитие Альпийского складчатого пояса. В палеогене и неогене поднялись горные цепи Атласа, Пиренеев, Альп, Апеннин, Балкан, Карпат, Крыма, Кавказа, Копетдага, Памира и Гималаев, завершилось формирование Анд, Кордильер и ряда других горных систем. Неоген характеризуется высоким положением материков (геократическая эпоха), чему соответствовала наибольшая степень осушения их территорий за весь фанерозой (с этим связано указанное выше преобладание открытых сухих ландшафтов в неогене).
Первый обширный ледниковый щит сформировался в Антарктиде в миоценовую эпоху неогена (около 20 млн. лет назад). До этого (в течение палеогена) в Антарктиде сохранялся умеренный прохладный климат и ледники имелись лишь в горах.
Наконец, в плейстоценовой эпохе четвертичного периода (около 3, а по некоторым данным даже 5 млн. лет назад) началось великое оледенение, сопровождавшееся развитием гигантских ледниковых щитов в Европе и Северной Америке. Оледенение прерывалось несколько раз резко наступавшими потеплениями (межледниковые эпохи, или интерстадиалы), во время которых ледниковый покров на континентах Европы и Северной Америки стаивал практически полностью. Затем следовало новое понижение температур с последующим оледенением. Период этих изменений составлял в среднем около 100 000 лет.
Максимальное из плейстоценовых оледенений (около 250 000 лет назад) покрывало территорию около 45 000 000 км2 (т. е. примерно 30% суши), ледниковый щит в Европе достигал широты 48°30', а в Северной Америке 37°. Объем льдов суши в это время составляя около 55 000 000 км3. Соответственно, уровень Мирового океана в эпохи оледенений значительно падал (на 85—120 м), и возникали сухопутные связи между разобщенными ныне проливами континентами и островами. Например, берингийская суша связывала в эпохи оледенений Аляску и Чукотку, Британские острова соединялись с Европой и т. д. Последнее оледенение завершилось около 10 000 лет назад.
РАЗВИТИЕ ЖИЗНИ В ПАЛЕОГЕНЕ
В начале палеоценовой эпохи фауна млекопитающих оставалась, по существу, той же, что и в позднемеловое время. В ней были представлены лишь группы, возникшие еще в мезозое: растительноядные многобугорчатые, внешне напоминавшие грызунов, но родственные прототериям — однопроходным, а также архаические представители сумчатых и плацентарных, питавшиеся насекомыми и другой мелкой добычей. Для всех архаических млекопитающих были характерны такие примитивные особенности, как относительно небольшой мозг, простые треугольные зубы (за исключением многобугорчатых), пятипалые конечности, опиравшиеся при передвижении на всю кисть и стопу (плантиградность). К середине палеоценовой эпохи разнообразие млекопитающих значительно возросло (настолько, что можно предположить начало расхождения некоторых предковых линий еще до конца мелового периода). Но основная адаптивная радиация плацентарных и сумчатых происходила в палеоцене и эоцене, когда сложились все основные отряды кайнозойскихмлекопитающих.
… но особенно замечательны были гигантские носороги (Indricotheriidae, рис. 62), распространенные в Азии в олигоцене и начале миоцена. Это были крупнейшие из известных науке наземных млекопитающих, превосходившие даже крупных хоботных (и уступавшие по длине и массе тела лишь завроподам); длина их тела достигала 7—8 м, высота в холке — до 5,5 м. Довольно длинная и мощная шея могла поднять огромную голову (череп имел длину до 1,2 м) на высоту около 8 м над землей. Гигантские носороги обитали в открытых ландшафтах, лесостепях и саваннах, которые были широко распространены в Азии уже в олигоцене (Б. А. Трофимов, В. Ю. Решетов, 1975). Вероятно, эти животные питались листвой деревьев с высокими кронами. Дальнейшая аридизация азиатских степей привела в неогене к вымиранию этих удивительных животных.
Между птицами и млекопитающими произошел своего рода «раздел сфер влияния». Большинство видов зверей, возникших от сумеречных и ночных мезозойских форм (см. выше), сохранило и в кайнозое предпочтение к ночной активности, тогда как птицы, потомки гелиотермных рептилий, остались преимущественно дневными животными и поныне. Ш. Д. Пьер (1975) подсчитал, что в составе современной фауны тропических лесов Габона и Панамы дневной образ жизни ведет 96% видов птиц, тогда как для 70% видов млекопитающих характерна ночная активность.
РАЗВИТИЕ ЖИЗНИ В НЕОГЕНЕ
Как мы уже упоминали,в течение неогена произошло значительное повышение материков (геократическая фаза), сопровождавшееся морской регрессией, иссушением климата и прогрессирующим развитием открытых ландшафтов. Одновременно происходило постепенное понижение средних температур на поверхности Земли. Эти изменения природных условий оказали глубокое влияние на флору, о чем уже говорилось выше, и фауну. В неогене получили преобладание виды наземных животных, приспособленные к жизни в открытых и относительно сухих местообитаниях типа лесостепей, степей и саванн. В олигоцене вымерли многие формы, тяготевшие к лесам, влажным лугам, заболоченным участкам и тому подобным биотопам, характерным для палеогена. Эта судьба постигла многие группы непарнокопытных. Зато парнокопытные начиная с олигоцена широко распространились и быстро прогрессировали. Особенно это относится к наиболее новым их группам: оленям (Cervidae), жирафам (Giraffidae), вилорогам (Antilocapridae) и многочисленным полорогим (Bovidae). Успех этих форм прежде всего связан со специализацией зубов и пищеварительной системы к переработке грубых растительных кормов (злаков и других растений степей и саванн), а также с приспособлениями к быстрому бегу по относительно плотной почве травянистых равнин.
ЧЕТВЕРТИЧНЫЙ ПЕРИОД
В плейстоцене происходило дальнейшее похолодание климата, сопровождавшееся постепенным отступанием теплолюбивой флоры и фауны в Евразии и Северной Америке к югу. Широкое распространение получили листопадные и хвойные леса, общий облик которых уже напоминал современные леса умеренного пояса. Состав фауны также приближался к современному; появились многие ныне существующие роды млекопитающих. Однако в плейстоцене сохранялись и более древние формы и группы животных (мегатерии, глиптодонты, макраухении, токсодонты, мастодонты, динотерии, саблезубы и др.), особенно многочисленные в Америке. Появились и некоторые своеобразные виды, которым не суждено было пережить плейстоценовую эпоху
Для всех видов животных мамонтовой фауны характерны приспособления к жизни в условиях низких температур, в частности длинная и густая шерсть. Густой и очень длинной рыжей шерстью (с длиной волос до 70—80 см) был покрыт и мамонт (Mammonteus, рис. 73) — северный слон, обитавший еще около 12 000 лет назад на огромных территориях Европы (достигая в отдельные эпохи к югу Италии, Крыма и Закавказья), Азии и Северной Америки.
Вымирание мамонтовой фауны отчетливо коррелирует с концом последнего оледенения 10—12 тыс. лет назад. Потепление климата и таяние ледников резко изменили природную обстановку в прежнем поясе приледниковой тундростепи; значительно возросла влажность воздуха и усилилось выпадение осадков; как следствие, на больших территориях развилась заболоченность, зимой увеличилась высота снежного покрова. Животные мамонтовой фауны, хорошо защищенные от сухого холода и способные добывать себе пищу на просторах тундростепи в малоснежные зимы ледниковой эпохи, оказались в крайне неблагоприятной для них экологической обстановке. Обилие снега зимой сделало невозможным добывание пищи в достаточном количестве. Летом же высокая влажность и заболачивание почвы, крайне неблагоприятные и сами по себе, сопровождались колоссальным возрастанием численности кровососущих насекомых (гнуса, столь обильного в современной тундре), укусы которых изнуряли животных, не давая им спокойно кормиться (как это происходит и ныне с северными оленями). Таким образом, мамонтовая фауна оказалась за очень короткий срок (таяние ледников происходило очень быстро, см. выше) перед лицом резких изменений среды обитания, к которым большинство составлявших ее видов не сумело столь быстро приспособиться, и она как целое прекратила существование.
В конце плейстоцена произошло еще одно значительное изменение фауны, правда, ограниченное масштабами Америки, но остающееся до сих пор загадочным. В Северной и Южной Америке вымерло подавляющее большинство крупных животных, столь обильных там прежде: и представители мамонтовой фауны, и обитавшие в более южных районах, где не было никакого оледенения, мастодонты и слоны, все лошади и большинство верблюдов, мегатерии и глиптодонты. Видимо, еще в плиоцене исчезли носороги. Из крупных млекопитающих сохранились лишь олени и бизоны в Северной Америке и ламы и тапиры в Южной. Это тем более удивительно, что Северная Америка была родиной и центром эволюции лошадей и верблюдов, сохранившихся до нашего времени в Старом Свете. Нет никаких признаков значительных изменений условий существования в конце плейстоцена на большей части территории Америки, не подвергавшейся оледенению. Более того, после появления в Америке европейцев, некоторые из завезенных ими лошадей оказались на воле, одичали и дали начало мустангам, быстро размножившимся в североамериканских прериях и южноамериканской пампе, условия которых оказались вполне благоприятными для этих животных.
После завершения последнего оледенения 10—12 тыс. лет назад Земля вступила в голоценовую эпоху четвертичного периода, в течение которой установился современный облик фауны и флоры. Условия жизни на Земле ныне значительно более суровы, чем на протяжении мезозоя, палеогена и большей части неогена. И богатство и разнообразие мира организмов в наше время, судя по всему, существенно ниже, чем за многие прошлые эпохи. В голоцене все большую возможность воздействия на среду обитания приобретает человек. В наше время с развитием технической цивилизации деятельность людей стала поистине важнейшим глобальным фактором, активно изменяющим биосферу.
ГЛАВА 6. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Человек относится к отряду приматов (Primates), и эволюционная история человека есть часть филогенеза этой группы. Поэтому рассказ о происхождении человека целесообразно начать с анализа общих признаков приматов, среди которых можно найти и предпосылки для развития особенностей строения, физиологии и поведения человека. Приматов можно кратко охарактеризовать как группу лесных теплолюбивых плацентарных млекопитающих, приспособившихся к лазающему, древесному образу жизни, сохранив достаточно примитивную основу организации.
РАННИЕ ЭТАПЫ ЭВОЛЮЦИИ ПРИМАТОВ
Приматы возникли от примитивных насекомоядных млекопитающих (Insectivora), отдельные группы которых перешли к лазающей, древесной жизни. К основанию эволюционного ствола приматов, возможно, близки древесные землеройки — тупайи (Tupaiidae), обитающие ныне во влажных тропических лесах Южной и Юго-Восточной Азии и прилежащих островов. Среди зоологов-систематиков до сих пор нет единства мнений в вопросе о положении тупайид в системе млекопитающих. Одни авторы рассматривают их как семейство отряда насекомоядных, другие помещают их среди приматов. Эволюционная ветвь насекомоядных, ведущая к приматам, обособилась, вероятно, еще до конца мезозоя. Из верхнемеловых отложений известны ископаемые остатки животных, которых ряд палеонтологов рассматривает уже в качестве примитивных приматов.
От каких-то примитивных лемуров примерно в середине палеогена возникли настоящие обезьяны — антропоиды (Anthropoidea). Вероятно, их обособление от лемуров было связано с переходом к дневной активности, сопровождавшимся усилением роли зрения, увеличением размеров и совершенствованием строения головного мозга, развитием стайного образа жизни и связанных с ним социальных форм поведения.
Древнейшие остатки антропоидов известны из нижнеолигоценовых отложений Египта. Однако антропоиды возникли, вероятно, еще раньше (может быть, в конце эоцена), поскольку в раннем олигоцене существовали уже достаточно разнообразные их представители.
ЭВОЛЮЦИЯ ЧЕЛОВЕКООБРАЗНЫХ ПРИМАТОВ
Расхождение филогенетических стволов, ведущих к двум семействам антропоморфов, т. е. человекообразным обезьянам (понгидам) и людям (гоминидам), произошло, возможно, уже в миоцене (по разным оценкам, от 15 до 25 млн. лет назад). Из верхнемиоценовых отложений Европы известны ископаемые остатки обезьян дриопитеков (Dryopithecus), которые по основным особенностям строения и размерам тела напоминали современных шимпанзе. Многие антропологи рассматривают дриопитеков как возможных непосредственных предков всех высших антропоморфов, т. е. понгид и гоминид. В неогене антропоморфные приматы достигли высшего расцвета. Их ископаемые остатки широко представлены в местонахождениях гиппарионовой фауны. Это говорит о связи миоплиоценовых антропоморфов с характерными для этой фауны открытыми ландшафтами (тогда как большинство приматов, и особенно низшие их представители, являются обитателями тропических лесов, столь обильных в палеогене). Вероятно, древнейшие человекообразные обезьяны обитали в разреженных лесах на границах лесных массивов и лесостепей.
В качестве наиболее древнего и примитивного представителя эволюционной линии гоминид многие антропологи рассматривают рамапитека (Ramapithecus), ископаемые остатки которого были обнаружены в верхнемиоценовых отложениях Индии, абсолютный возраст которых определен в 12 млн. лет, а также в близких по времени возникновения отложениях в Кении и Венгрии (Р. Лики, 1976). К сожалению, пока еще нет палеонтологических данных о ближайших последующих этапах филогенеза гоминид. Зато большое количество ископаемых остатков высших антропоморфов известно начиная с верхнеплиоценовыхотложений, имеющих возраст от 3,5 млн. лет и моложе. Эти остатки принадлежат различным формам африканских австралопитеков (Australopithecinae): австралопитека (Australopithecus), парантропа (Paranthropus), плезиантропа (Р1еsianthropus), зинджантропа (Zindjanthropus). Впервые остатки австралопитеков были обнаружены в Южной Африке в 1924 г. С тех пор эти антропоморфы остаются в центре внимания антропологов и являются предметом оживленной дискуссии.
Австралопитеки, обитавшие в Южной и Восточной Африке во второй половине плиоцена и начале плейстоцена (от 3,5, а по некоторым данным от 5 млн. лет до 1 млн. лет назад), во многих отношениях были гораздо ближе к людям, чем любые человекообразные обезьяны. Судя по строению их таза и скелета задних конечностей, австралопитеки постоянно использовали двуногое передвижение (хотя их бипедальность была значительно менее совершенной, чем у современных людей). Пропорции их черепа (рис. 74) и особенности зубной системы также ближе к соответствующим показателям гоминид, чем понгид. Средний объем мозговой коробки австралопитеков, однако, оставался близким к таковому человекообразных обезьян (522 см, достигая в отдельных случаях 650 см).
Судя по этим данным, типичные плейстоценовые австралопитеки действительно существовали одновременно с более прогрессивными гоминидами и поэтому не могут рассматриваться как предки последних. Однако остается вполне вероятным общее происхождение всех плейстоценовых гоминид (включая австралопитеков) от каких-то позднеплиоценовых предков, которыми вполне могли быть более ранние австралопитековые, пока еще мало известные. Во всяком случае, в морфологическом отношении австралопитеки занимают промежуточное положение между высшими гоминидами и человекообразными обезьянами (понгидами) и дают представление о первом важном этапе на пути гоминизации.
Что же касается вопроса, рассматривать ли австралопитеков как высших человекообразных обезьян или как наиболее примитивных людей, очевидно, решить его удалось бы при наличии четких критериев, определяющих принадлежность к людям. В качестве таковых указывались: постоянное использование двуногого передвижения, выпрямленное положение тела, увеличение первого пальца руки со сложной дифференциацией его мышц, увеличение головного мозга и относительных размеров больших полушарий, некоторые особенности зубной системы. По одним из этих показателей австралопитеки ближе к людям, по другим — к человекообразным обезьянам, занимая в целом промежуточное положение между теми и другими.
Трудовая деятельность стала важнейшим фактором дальнейшей эволюции человека. Поскольку использование орудий при высокоразвитой социальности (начинавшей формироваться еще у низших антропоидов) давало человеку огромные преимущества и позволяло осваивать новые местообитания и новые природные ресурсы, естественный отбор благоприятствовал таким изменениям организации древних людей, которые позволяли совершенствовать трудовую деятельность и социальное поведение. Это были в первую очередь прогрессивные изменения структуры и массы головного мозга, а также мускулатуры и скелета передних конечностей, с развитием механизмов тонкой нервной координации движений. Далее, отбор благоприятствовал развитию у людей таких форм поведения, которые облегчали их общение в процессе трудовой деятельности, в организации совместной охоты или защиты от нападений хищников. В связи с этим должны были совершенствоваться способы обмена информацией в частности система звуковой сигнализации, развитие которой привело к формированию членораздельной речи, а на ее основе — второй сигнальной системы, сыгравшей столь огромную роль в формировании человеческого интеллекта и культуры.
ЭВОЛЮЦИЯ РОДА HOMO
Филогенетическую историю гоминид, в ходе которой постепенно формировались особенности современного человека, разделяют на ряд последовательных стадий;
1) «предшественники людей», или протоантропы; 2) архантропы; 3) палеоантропы; 4) неоантропы. Протоантропы были представлены австралопитеками, существовавшими в позднем плиоцене и плейстоцене (5,5—1,0 млн. лет назад, но, возможно, возникли еще раньше). Австралопитеки создали костяную и галечную олдовайскую культуру.
Архантропы известны по многочисленным находкам в самых различных уголках Старого Света. Первая из этих находок была сделана еще в 1891 г. Е. Дюбуа, обнаружившим на острове Ява ископаемые остатки существа, получившего название питекантроп или обезьяночеловек. Первоначально разные архантропы рассматривались как представители разных родов: питекантропы с Явы, синантропы из ряда местонахождений на территории Китая, гейдельбергский человек в Европе, атлантропы в Северной Африке и др. Ныне большинство исследователей считают всех архантропов принадлежащими к одному виду Homo erectus (человек выпрямленный), в пределах которого выделяют до 9 подвидовых форм. До недавнего времени считали, что архантропы существовали на Земле в среднем плейстоцене, со времени гюнцского и до миндельского оледенения (т. е. 700 000—300 000 лет назад), однако новые данные (например, упомянутые выше находки, сделанные Р. Лики на восточном берегу озера Рудольф) и уточнение датировок расширяют время существования архантропов до огромного диапазона (2,6—0,2 млн. лет назад).
Следующая стадия в эволюции гоминид, палеоантропы, представлена так называемыми неандертальцами (Homo neanderthalensis), видовое название которых связано с первой находкой ископаемых остатков этих людей в долине Неандерталь вблизи Дюссельдорфа. Неандертальцы, как и архантропы, были распространены практически по всей территории Старого Света и очень разнообразны. Они появились на Земле примерно 300 000 лет назад (во время миндель-рисского межледниковья) и просуществовали до первой половины вюрмского оледенения, т. е. примерно до 35 000 лет назад
Палеоантропами была создана культура среднего палеолита. Неандертальцы хоронили своих умерших с определенными погребальными обрядами, что позволяет предполагать наличие у них достаточно развитого абстрактного мышления (Арамбур, 1966).
Стадия неоантропов соответствует человеку современного вида (человек разумный — Homo sapiens). Древнейшие неоантропы, называемые кроманьонцами (по месту первой находки их ископаемых остатков в 1868 г. в гроте Кро-Маньон, на территории французской провинции Дордонь), известны начиная со времени среднего вюрмского оледенения в позднем плейстоцене (около 38 000 — 40 000 лет назад). Кроманьонцы (рис. 78) уже вполне соответствовали антропологическому типу современного человека, отличаясь лишь незначительными особенностями: несколько менее высокий свод черепа, сильнее развитая зубная система и др.
Кроманьонцы были создателями культуры позднего палеолита, характеризующейся высоким совершенством обработки камня и кости. Именно кроманьонцы были творцами пещерных рисунков, запечатлевших животных мамонтовой фауны, а также древнейших скульптурных изображений и первых музыкальных инструментов. Можно на этом основании утверждать, что с неоантропами возникает искусство
На этих основаниях теория полицентризма считает более вероятной параллельную эволюцию нескольких филетических линий гоминид, с независимым приобретением особенностей последующих стадий, от архантропов до неоантропов на разных континентах Старого Света. Современные большие человеческие расы (европеоиды, негроиды, монголоиды и австралоиды) выводятся при этом от разных подвидов палеоантропов и даже архантропов. В Новый Свет человек проник относительно недавно (около 25 000—30 000 лет назад) и, как предполагают, уже на стадии неоантропа.
С позиций моноцентризма современные человеческие расы представляют собой относительно поздно (25 000—35 000 лет назад) сформировавшиеся группировки, обособившиеся в процессе расселения неоантропов из области их возникновения.
ххххххххх
-- А вот статья из википедии, где даны сведения о доисторическом периоде человечества.
https://ru.wikipedia.org/wiki/Первобытное_общество
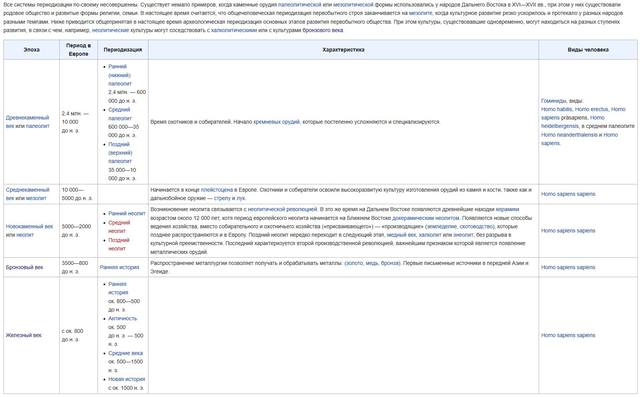
-- Взяв из нее сроки указанные историками, я добавил цифры из "Тайной Доктрины" ЕПБ. Получилась такая таблица. Возможно при этом допустил небольшие неточности. Тогда чуть позже их исправлю. (Размер картинки можно увеличить щелчком мыши по ней)

ххххххххххххххх

хххх
-- Из ролика о происхождении человека мы видим, насколько нелепо выглядит версия ученых по сравнению с той, что дана Великими Учителями.
Происхождение человека. Антропогенез.
хххххххх
-- И в завершении раздела о версии истории планеты Земля от УЧЕНЫХ размещу видеовариант.
ИСТОРИЯ, УХОДЯЩАЯ В ГЛУБЬ ВРЕМЕН 1, 2, и 3-я серия
хххххх
Смитсоновский институт признал уничтожение тысяч скелетов гигантов
https://u.to/FHK0Fg
хххххххх
|
| |
|
|
| kualspb_2013 | Дата: Пятница, 15.11.2019, 18:56 | Сообщение # 4 |
|
Генералиссимус
Группа: Администраторы
Сообщений: 2147
Статус: Offline
| _004
-- Заканчивая изложение истории Земли в версии НАУКИ, хотелось бы добавить, что в дарвиновской теории происхождения человека разумного за последнее столетие обнаружено огромное количество ошибок и противоречивых утверждений. Часть из них была опубликована для общего доступа еще в конце 19 века во втором томе "Тайной Доктрины". Там приводится очень много фактов, с которыми ученым стоило бы внимательно ознакомиться. Но и без этих данных в 20 веке было открыто много новых фактов, доказывающих, что случайным образом (под воздействием слепых сил природы) ни человек, ни даже простейшие микроорганизмы и растения не могли появиться ни на Земле, ни в каком другом месте Вселенной.
Прежде, чем читатель этого сайта приступит к знакомству с темой об истории планеты в версии Махатм, (изложенной в "Тайной Доктрине"), необходимо пояснить, что она была написана ТОЛЬКО ПОСЛЕ ТОГО, как Махатмы вступили в переписку с двумя образованными европейцами того времени (через посредство Е.П.Блавасткой) и в результате появились книги одного из них (А.П.Синнет) под названием "Письма Махатм", "Эзотерический буддизм" и некоторые другие. И первая УПРОЩЕННАЯ ВЕРСИЯ от Великих Учителей истории нашей планеты была написана в книге "Эзотерический буддизм". На вопросы Синнета отвечали сами Махатмы. И поэтому ниже будет размещена информация из этой книги.
Только сразу уточню, что это именно упрощенная версия. И в ней содержалось много ОШИБОК. Часть из них была устранена в "Тайной Доктрине". Но и в последней содержалось все же много упрощений, которые в головах читателей превратились в ОШИБКИ. И хотя в дальнейшем эти упрощения были заменены в последующих УЧЕНИЯХ от Махатм более правильной информацией, (например в учении Макса Генделя), все же и эти учения в свою очередь тоже содержали УПРОЩЕНИЯ. И их каждый раз надо было исправлять. В этой теме мы постараемся проследить НЕСКОЛЬКО ТАКИХ попыток Махатм дать более полную и точную информацию на тему истории человечества. Но сделать это будет не просто по следующим причинам. Великие Учителя не хотели бы, чтобы в головах любого интеллектуально развитого читателя при прочтении книг, записанных их учениками, возникла ЧЕТКАЯ, ТОЧНАЯ и БЕЗОШИБОЧНАЯ картина эволюции на нашей планете. Дается только то, что СООТВЕТСТВУЕТ нашему уровню развития. Какие-то вещи нам ПОКА объяснять ОПАСНО и они оставляются не раскрытыми для тех, кто не достиг ступени ПРИНЯТОГО УЧЕНИЧЕСТВА. Это очень высокий духовный уровень, при котором ученик уже не способен нанести вред ПЛАНАМ Иерархии Света. Те же, кто не достиг такого уровня, при получении некоторых сокровенных знаний могут свернуть на путь называемый в оккультизме Левой Тропой -- путь черной магии и эгоизма.
ЦИТАТА (Эзотерический буддизм", Глава VII) : Изучающие оккультную философию, приступая к рассмотрению этого вопроса с умом, уже обильно оснащённым в других направлениях, склонны с самого начала делать не совсем правильные выводы. Сразу все объяснить невозможно, а самые первые, обобщенные объяснения нередко заставляют наиболее вдумчивых и умственно активных читателей домысливать совершенно неверные подробности. Такие читатели не хотят довольствоваться лишь общими контурами даже временно. Их воображение стремится заполнить все пустоты, и если дать ему возможность некоторое время трудиться без направляющего вмешательства извне, то впоследствии обладатель этого живого воображения с большим удивлением заметит, что продолжающая поступать к нему информация на ту же самую тему не согласуется с тем, чему его (как ему самому кажется) учили раньше. Автор этого трактата прилагал все усилия к тому, чтобы способ его изложения не допускал поспешного роста сорняков недопонимания в умах читателей, однако сами эти усилия иногда требуют забегать вперёд, набрасывая общие контуры и выпуская некоторые детали, нередко довольно важные, оставляя их до повторного, более подробного рассмотрения.
-- Так и происходило исторически в последнее столетие. Ошибки Синнета чуть позже исправила ЕПБ, написав "Тайную Доктрину". Теософы превратили ее учение в ДОГМУ и многие из них не стали учитывать те ИСПРАВЛЕНИЯ и ДОПОЛНЕНИЯ, которые появились в последующих учениях от Махатм. В данной теме мы постараемся начать развязывать этот клубок противоречий, накопившийся из-за того, что разрозненные части ЕДИНОГО УЧЕНИЯ, пока не связаны ЕДИНОЙ НИТЬЮ последовательного изучения РАЗНЫХ учений. Ученикам пока легче изучить только одно какое-то учение. И на основании этих знаний сделать вывод, что дальше следовали лишь ИСКАЖЕНИЯ тех основ, которые были усвоены из источника со 100% надежностью в том, что информация получена от Махатм.
хххххххххххххххххххххххх
-- Должен предупредить, что в истории Земли ПОКА у ученых есть не мало "БЕЛЫХ ПЯТЕН". И есть много находок археологов, которые не нашли ПОКА внятных объяснений со стороны науки. Приведу несколько примеров.
Саксайуаман- Вес каменных глыб нижнего яруса составляет несколько сотен тонн, вес самого большого блока, достигающего более 8 метров в высоту, оценивается примерно в 3500 тонн. Огромные блоки из андезита – очень твердого и сложного в обработке материала, образуют полигональную кладку с подгонкой камней друг к другу без какого-либо раствора https://u.to/FS1bFg
Больше всего, в Саксайуаман поражает кладка зигзагообразных стен, особенно в нижнем ярусе, который сложен из самых больших блоков. Видимо создателям этих стен перемещение монолитов в сотни тонн весом и обработка твердого андезита не составляли абсолютно никаких проблем. Сложно представить, при помощи каких технологий, и каким инструментом блоки из твердого андезита обрабатывались так, что достигалось идеальное сопряжение стыков. Некоторые поверхности блоков даже отшлифованы. Саксайуаман является самым грандиозным, но далеко не единственным памятником с подобными циклопическими постройками. В так называемой «царской долине», в которой расположен Куско сохранились и другие памятники с огромными мегалитическими постройками – Мачу Пикчу, Олльянтайтамбо и другие. В 2003 году испанский археолог Ансельм Пи Рамба обнаружил подземный тоннель, соединявший Саксайуаман с центром Куско. Тоннель проложен на глубине около ста метров от поверхности и имеет протяженность около 2 км. Гарсиласо де ла Вега писал о целом подземном городе под инкской столицей, который состоял из целого лабиринта галерей, подземных храмов и хранилищ.
 х х 

хххххххххххх
Самым знаменитым сооружением древнего Баальбека, пожалуй, является храм Юпитера, который был построен в I веке нашей эры.
https://u.to/Xy9bFg
От грандиозного храма, который являлся самым большим в Римской империи, осталось каменное основание и шесть колонн. Причем в основании храма имеются невероятных размеров три каменных блока, которые получили название трилитон Баальбека. По оценкам специалистов, масса этих известняковых блоков составляет 300-800 тонн, что намного превышает вес блоков, из которых сложена пирамида Хеопса в Египте.
 х х 
хххххххххх
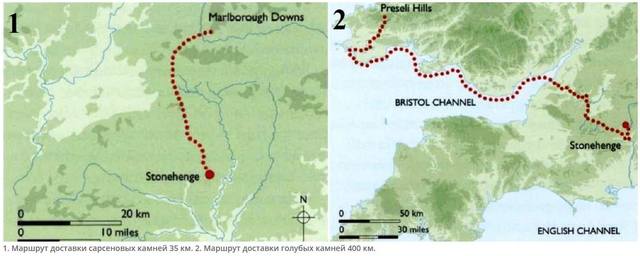
Архитектура древних - Стоунхендж. https://u.to/4TFbFg
Трилиты образуют подкову диаметром 15 метров, открытую в сторону аллеи, они установлены симметрично: самая маленькая пара трилитов была 6 метров высотой, следующая пара 6,5 метра, а самым большим был единственный центральный трилит высотой в 7,3 метра. Самые огромные мегалиты из центральной пары трилитов весят до 50 тонн каждый… И напоследок вся центральная часть была «окольцована» тридцатью огромными сарсеновыми блоками, высота 5 метров (4,1 м. над землей), 2 метра ширина, толщиной в 1,1 метр и весом 25 тонн, каждый. Диаметр «кольца» составлял 29,6 метра, расстояние между мегалитами 1 метр… На этих опорных блоках сверху лежали поперечные блоки, крепились они с помощью шипов и пазов. Длина перемычек составляла около 3,2 метра, ширина 1 метр и толщина 0,8 метра и весом до 6 тонн каждый, круг из перемычек был выровнен в плоскости, так что верх каждой перемычки находился на высоте 4,9 метра над землей.
 х х 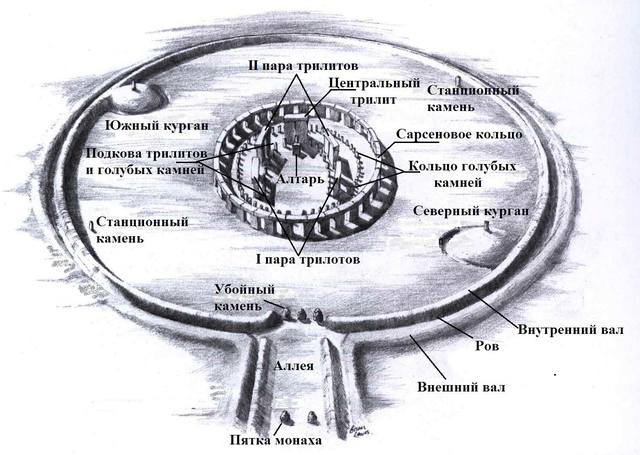
хххххххххх
 х х 
Дольмен Sorginetxe в Arrizala и дольмен в Южной Корее
 х х 

Галерея Эйвбери — культовый объект из мегалитических гробниц и святилищ. Находится в графстве Уилтшир, в Англии.

Доль-де-Бретань менгир во Франции
ххххххххххх
-- Махатмы утверждают, что остров Пасхи -- это остатки Лемурийской цивилизации, которая существовала несколько миллионов лет назад. Тогда на Земле рост людей был намного больше, чем сейчас. И в мифах народов мира они упоминаются, как ВЕЛИКАНЫ.
 х х 
 х х 
-- А вот версия ученых. По моему это просто смешно продолжать верить в такие ГИПОТЕЗЫ.

-- Все фото увеличиваются при щелчке по ним мышкой.
хххххххххххх
Неожиданная правда об острове Пасха.С каким умыслом это сделано.Территория загадок
Письменность острова Пасхи (рассказывает историк Альберт Давлетшин)
ОСТРОВ ПАСХИ - НОВЫЕ НАХОДКИ и ВОПРОСЫ/ Часть #1
ххххххххххх
5 ОТКРЫТИЙ, ДОКАЗЫВАЮЩИХ СУЩЕСТВОВАНИЕ ВЕЛИКАНОВ
Антинаучная гипотеза неожиданно нашла подтверждение,это потрясло научный мир.НОВАЯ теория ЭВОЛЮЦИИ
ххххххххх
Найден 10 – ти метровый человек-гигант который перевернул всю историю человечества
ххххххх
-- Беседа с учеными о том нужно ли воевать с лженаукой и что таковой считать.
Большое интервью - Ученые НЕ против и Инквизиция XXI века
ххххххх
|
| |
|
|
| kualspb_2013 | Дата: Среда, 17.01.2024, 23:53 | Сообщение # 5 |
|
Генералиссимус
Группа: Администраторы
Сообщений: 2147
Статус: Offline
| хххххххххх
Доисторическая Земля. Путешествие в прошлое
ххххххххххх
|
| |
|
|





